
Spider monkeys have a well-developed olfactory sensitivity for mold-associated odorants
Other studies using the same experimental procedures have shown a well-developed olfactory sensitivity for other odorant classes in this primate species, too. The findings of the present study do not support the long-held belief that primates have a poorly developed sense of smell.
Interindividual variability was generally low, varying at the most by a dilution factor of 100 for trans-2-octen-1-ol, which in turn is smaller than the range of interindividual variability reported in studies on human olfactory sensitivity. For 1-octen-3-ol all three individuals even reached the same detection threshold value.
Even though no proper statistical comparisons can be drawn due to the small number of individuals used, I examined between-odorant differences (based on n=3 data points) by considering whether the ranges of threshold values overlap or not, in order to at least get a first impression of possible differences in sensitivity.
Considerations about the molecular structural features by comparison of the threshold values for mold-associated odorants with other previously tested odorants
The olfactory detection threshold values determined in the present study ranged from 0.41 ppb for 3-octanone to 3.70 ppm for 2-methyl-1-propanol.
The lowest detection thresholds found here were generally below 1 ppm and correspond to the aliphatic compounds with an unbranched backbone chain of eight carbons. On the other hand, the odorants with a branched backbone of only four or five carbons used in this study were found to have detection thresholds higher than 1 ppm for all individuals, except Edgar and Kelly for 2-methyl-1-butanol and Kelly for 3-methyl-1-butanol. These findings lend further support to an existing correlation between backbone carbon chain length and detectability in spider monkeys, as found in earlier studies on aliphatic esters, aliphatic alcohols and aldehydes and putrefaction-associated odorants.
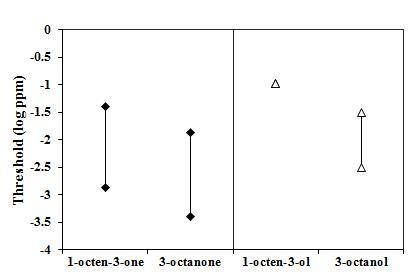
Presence or absence of double bonds and detectability
The odorant pairs 1-octen-3-one/3-octanone and 1-octen-3-ol/3-octanol (Figure 1) used in this study may be provide information about the effect of the presence or absence of a double bond in the molecule on detectability. The overlap of the detection threshold ranges of the first odorant pair indicates that this structural feature does not seem to affect detectability, as reported by Løtvedt and co-workers (2012). However, in the second odorant pair there does not seem to be an overlap of detection ranges, indicating a higher sensitivity to the substance lacking a double bond. Due to the inconclusive nature of the findings in the present study, more research should be conducted to unveil whether the presence of a double bond affects olfactory sensitivity in a systematic manner or whether some odorant classes may be an exception to the rule in spider monkeys
The odorants trans-2-octen-1-ol (tested here; detection range: 1.5 – 152 ppb) and 1-octanol (Laska et al. 2006a; detection threshold value: 4.8 ppb) differ in the presence or absence of a double bond in the carbon backbone chain. Here, too, no difference in detectability is apparent, further suggesting that the presence or absence of a double bond may not affect olfactory sensitivity in spider monkeys.
Type of functional group and detectability
The olfactory detection thresholds for 3-octanone (tested in this study) ranged from 0.41 ppb to 4.1 ppb, while the range of the threshold values for 2-octanone (Eliasson et al. 2015) reached as low as 0.034 ppm to as high as 0.34 ppm. Here, though, there is a remarkable difference in terms of the olfactory sensitivity of substances that differ in their position of the functional group, i.e., spider monkeys are more sensitive to 3-octanone than to 2-octanone. This difference is rather interesting, posing an exception to previous findings on the effect of the position of the functional group on detectability. Some conclusions can be drawn from this discrepancy. The results obtained in the present study are reliable and comparable with previous ones, as the same method was used, as well as a low inter-individual variability was reported. The most straightforward explanation is that the position of the functional group may have an effect on detectability only in some odorant classes and may not be a general rule. Thus, future research should focus on this issue.
Branching of the carbon chain and detectability
Finally, further conclusions can be drawn by comparing the olfactory detection thresholds of structural isomers, i.e., molecules with the same atomic composition but having a different structural arrangement. The threshold values of the odorant 2-methyl-1-propanol (tested here) ranged between 0.56 ppm and 1.57 ppm, whilst the range of threshold values for the odorant 1-butanol (Laska et al. 2006a) was 0.26 – 0.86 ppm. There is a small overlap between these value ranges but the detection thresholds outside of the overlapping area indicate that spider monkeys are more sensitive to the unbranched odorant, 1-butanol, than to the branched one, 2-methyl-1-propanol. Also, the odorants 2-methyl-1-butanol (detection threshold range: 0.56 – 1.57 ppm) and 3-methyl-1-butanol (detection threshold range: 0.48 – 1.44 ppm), both tested in the present study, are branched structural isomers of the odorant 1-pentanol (Laska et al. 2006a; detection threshold range: 0.4 ppb – 0.04 ppm). Spider monkeys are clearly more sensitive, again, to the unbranched odorant 1-pentanol than to its branched isomers. These findings have not been reported in earlier studies, and indicate that another odor structure-activity relationship may be at play. Thus, future studies should systematically investigate the characteristics of this relationship.
Sensitivity towards mold-associated odorants vs. other odorant classes
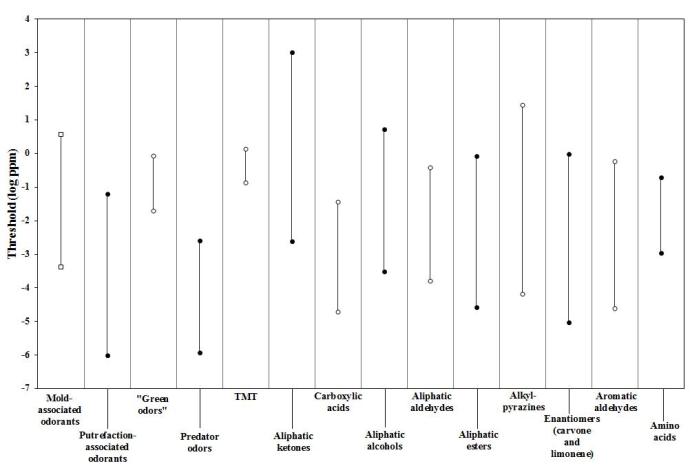
Figure 2 depicts a comparison of the detection threshold ranges for spider monkeys across the odorant groups available on the literature, including the ones used in the present study.
Considering the numerous studies conducted on the sensitivity of spider monkeys to different odorant classes, one may conclude that sensitivity for mold-associated odorants overlaps with the sensitivity for most of the other chemical classes tested so far in spider monkeys.
Sensitivity towards mold-associated odorants in other species
Across the available literature, olfactory detection thresholds for the odorants used in the present study have only been reported for humans.
Figure 3 shows the ranges of olfactory detection threshold values for both humans and spider monkeys for mold-associated odorants. Humans are clearly more sensitive to five of the eight mold-associated odorants tested in the present study than spider monkeys, whereas for three of the eight odorants, the human threshold range encompasses that of spider monkeys.
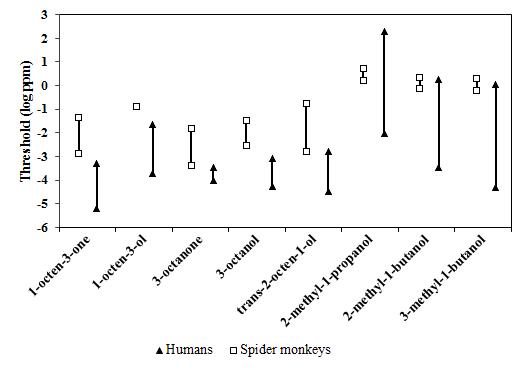
Several explanations for a difference in olfactory performance among different species have been proposed. Some authors suggested that the olfactory capabilities are correlated with the absolute or relative sizes of the olfactory bulbs. Humans have a higher absolute size of the main olfactory bulb (114 mm3) compared to spider monkeys (90.4 mm3). The same is not observed in terms of relative main olfactory bulb size, where spider monkeys surpass humans (0.9% vs. 0.09%, respectively). These findings indicate that olfactory bulb size does not explain a between-species difference in olfactory sensitivity, along with other findings found throughout the literature.
Other authors proposed that the number of functional olfactory receptor genes or the proportion of olfactory receptor pseudogenes are indicators of the olfactory sensitivity of a certain species. Spider monkeys have a higher number of functional olfactory receptor genes (≈900) than humans (≈396). If a clear correlation would exist, spider monkeys should be expected to be more sensitive to mold-associated odorants than humans, which is not the case. Once again, the lack of a correlation between genetic factors and olfactory sensitivity has also been reported in other studies, lending further support to the notion that genetic factors are not good predictors of olfactory performance.
Finally, the behavioral relevance of the odorants has been suggested to explain between-species differences in olfactory detection thresholds and increasing evidence supports this hypothesis. A species should be expected to be particularly sensitive to a certain odorant not only if it is present in its chemical environment, but also if it is relevant for its behavioral repertoire. For example, rats are markedly more sensitive to the odorant 2,4,5-trimethylthiazoline (TMT), found in the anal gland secretions of the red fox, than spider monkeys or pigtail macaques. As red foxes are known to predate rats, but not spider monkeys and pigtail macaques, it is, therefore, intuitive that rats are more adapted to detect this odorant at lower concentrations than the primate species.
Behavioral significance
Olfactory sensitivity seems to be better explained by the behavioral relevance of the odorants than by anatomical and genetic features, not only in a between-species comparison but also in a within-species comparison. Mold-associated odorants comprise a group of volatile substances produced by numerous strains of fungi. Fungal species are known to decrease nutritional content of the food material they develop in as well as to pose health hazards by means of mycotoxins and spores. Moreover, some substances (e.g. 1-octen-3-ol) are reported to be harmful at the tissue level in humans, rendering food-spoiling fungi potentially more hazardous than food-spoiling bacteria. Therefore, mold-associated odorants may work as cues of fruit edibility for the animals that depend on them, such as spider monkeys.
For many years it was implied that vision would be the predominant sense involved in the food selection process, as primates are traditionally viewed as “visual” animals. However, several studies documented that primates rely on other sensory information as well in the food selection context. For example, it has been reported that primates often smell, manipulate and lick food items in their natural habitat before consuming them. Even in captivity, studies with spider monkeys indicate that they rely on their senses of smell and touch to assess the quality of novel food and in the following inspections individuals tend to use their vision for familiar food items, though familiar food items may be still inspected by means of olfaction to check the quality of the fruit if other external cues, like color, do not clearly indicate so. This interplay of the senses in foraging and food selection indicates that there is valuable olfactory information that can be gathered from food prior to its consumption.
Also, the olfactory sensitivity of the spider monkeys to mold-associated odorants may be high enough to detect fungi-spoiled food when it is already contaminated with mycotoxins or even slightly before this stage, as there is a reported positive correlation between mycotoxin and volatile metabolites production. Furthermore, research indicates that the knowledge about the edibility of potential food needs to be learned and thus relying on various sensory information throughout that learning process is adaptive.
Future research should put extra effort in assessing the sensitivity to other mold-associated odorants referred in the literature, such as geosmin, a substance reported to have a mushy, earthy odor. Furthermore, other species of non-human primates and other non-primate mammals should be tested to shed light on the impact that the chemical environment has on the species’ olfactory capabilities.
Responsible for this page:
Director of undergraduate studies Biology
Last updated:
06/25/17